Module 5. La biodiversité
Introduction
Le terme biodiversité ![]() , un néologisme tiré de l’expression biological diversity, est devenu au fil du temps un mot à la mode pour divers acteurs dans notre société, que ce soit les groupes de pression environnementaux, les chefs d’entreprise ou les politiciens pour n’en nommer que quelques-uns. Cette pluralité d'utilisateurs fait en sorte qu’il existe plus de 80 définitions de ce terme (Spicer, 2006). La définition retenue dans ce module est celle de la Convention sur la diversité biologique
, un néologisme tiré de l’expression biological diversity, est devenu au fil du temps un mot à la mode pour divers acteurs dans notre société, que ce soit les groupes de pression environnementaux, les chefs d’entreprise ou les politiciens pour n’en nommer que quelques-uns. Cette pluralité d'utilisateurs fait en sorte qu’il existe plus de 80 définitions de ce terme (Spicer, 2006). La définition retenue dans ce module est celle de la Convention sur la diversité biologique ![]() :
:
« La variabilité des organismes vivants de toute origine y compris, entre autres, les écosystèmes terrestres, marins et autres écosystèmes aquatiques et les complexes écologiques dont ils font partie; cela comprend la diversité au sein des espèces et entre espèces ainsi que celle des écosystèmes. »
On remarque que la biodiversité doit être considérée à trois niveaux : la diversité des espèces, la diversité génétique ![]() et la diversité des écosystèmes. La diversité des espèces inclut toutes les espèces sur le globe, qu’elles soient unicellulaires (p. ex. les bactéries, les protozoaires) ou pluricellulaires (p. ex. les plantes, les animaux, les champignons). La diversité génétique, quant à elle, est la variété du matériel génétique des êtres vivants. Cette variété est étudiée à l’échelle d’une population ou de tous les individus de la même espèce. Enfin, la diversité des écosystèmes est celle que l’on observe dans une région donnée ou sur la planète (Primack, 2008). Certains auteurs ajoutent la diversité des populations d’une même espèce (chapitre 4 du volume 1 du MEA
et la diversité des écosystèmes. La diversité des espèces inclut toutes les espèces sur le globe, qu’elles soient unicellulaires (p. ex. les bactéries, les protozoaires) ou pluricellulaires (p. ex. les plantes, les animaux, les champignons). La diversité génétique, quant à elle, est la variété du matériel génétique des êtres vivants. Cette variété est étudiée à l’échelle d’une population ou de tous les individus de la même espèce. Enfin, la diversité des écosystèmes est celle que l’on observe dans une région donnée ou sur la planète (Primack, 2008). Certains auteurs ajoutent la diversité des populations d’une même espèce (chapitre 4 du volume 1 du MEA ![]() ).
).
5.1 Pourquoi s’intéresser à la biodiversité?
L’importance que l’on donne à la biodiversité va au-delà de la valeur sentimentale que plusieurs personnes lui accordent. En effet, la biodiversité favorise le fonctionnement normal des écosystèmes. La diversité des espèces fait en sorte, par exemple, que le cycle de l'azote demeure efficace et favorise la croissance normale des arbres et des plantes en rendant disponible l'azote emprisonné dans les végétaux morts. La diversité génétique, quant à elle, permet aux organismes de mieux résister et de survivre aux changements qui surviennent dans leur milieu (p. ex. les maladies, les changements climatiques). Enfin, la variété des écosystèmes contribue aux processus qui impliquent plusieurs écosystèmes. Par exemple, un couvert forestier limite l’érosion du sol, qui autrement entraînerait des sédiments dans les cours d’eau et dégraderait les habitats aquatiques (module 2 ![]() et module 3
et module 3 ![]() ).
).
La biodiversité a également des impacts majeurs sur les populations humaines :
- La diversité des espèces permet d’offrir les services auxquels nous sommes habitués. Ces services ne se limitent pas seulement à ceux présentés au module 1
 (p. ex. la nourriture, la filtration de l’eau). Ces services facilitent également la production de biens et de services. C’est le cas notamment en agriculture ou en foresterie où les populations d’insectes ravageurs sont contrôlées par diverses espèces d’oiseaux et d’insectes.
(p. ex. la nourriture, la filtration de l’eau). Ces services facilitent également la production de biens et de services. C’est le cas notamment en agriculture ou en foresterie où les populations d’insectes ravageurs sont contrôlées par diverses espèces d’oiseaux et d’insectes.
- La diversité génétique permet, quant à elle, d’améliorer les services que nous tirons des êtres vivants. Par exemple, la diversité génétique présente chez le blé permet la création de variétés de blé qui démontrent une productivité supérieure ou une résistance particulière aux maladies.
- La diversité des écosystèmes assure les besoins en nourriture et en habitats d’un plus grand nombre d’espèces (Primack, 2008).
- La diversité des populations fait en sorte que les services écologiques sont possibles sur une plus grande partie du territoire. Une telle diversité favorise par ailleurs une plus grande diversité génétique des espèces (chapitre 4 du volume 1 du MEA ).
5.2 Les concepts de base de la biodiversité
La compréhension des enjeux sur la biodiversité implique la connaissance de concepts particuliers à l’écologie. Les concepts abordés sont les suivants :
- L'évolution de la vie et de la diversité;
- La population;
- Les fonctions dans les écosystèmes.
5.2.1 L'évolution de la vie et de la diversité
La Terre a fait son apparition il y a de 3,5 à 4,5 milliards d’années (figure 5.1). Ce n’est que 1 à 2 milliards d’années après sa formation que les premières formes de vie, les procaryotes (p. ex. les bactéries), sont apparues. La présence des premiers animaux pluricellulaires a pu être évaluée à 1,5 milliard d’années avant aujourd’hui, suivi des premiers animaux il y a 700 millions d’années. Ces animaux étaient des invertébrés marins sans exosquelette tels que les éponges et les méduses. Deux cents millions d’années se sont ensuite écoulées avant que les vertébrés et les plantes fassent leur apparition. Les assemblages de mammifères, d’oiseaux et de plantes que nous connaissons aujourd’hui sont apparus au cours des derniers 70 millions d’années alors que les humains ont foulé le sol pour la première fois il y a 100 000 à 200 000 années (chapitre 4 du volume 1 du MEA ![]() ).
).
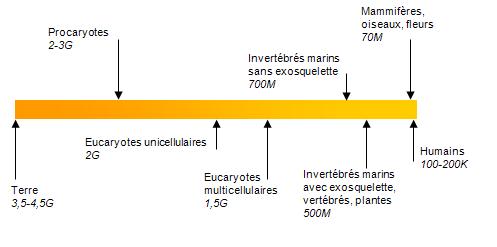
Figure 5.1 Ordre d’apparition des différentes formes de vie depuis la formation de la Terre. (Les périodes sont en années avant aujourd’hui. G = milliard d’années; M = million d’années; K = milliers d’années.)
Les biologistes de l’évolution croient que les espèces que nous connaissons aujourd’hui proviennent d’une seule et unique espèce. Les situations qui peuvent mener à l’apparition de nouvelles espèces sont les suivantes :
- Un groupe d’individus isolés géographiquement (p. ex. les individus sur une île très éloignée du continent - spéciation allopatrique );
- Un groupe d’individus qui ne sont pas isolés géographiquement, mais qui sont présents dans des milieux différents (p. ex. les plantes dans des milieux humides vs secs - spéciation parapatrique);
- Un groupe d’individus qui ne sont pas isolés géographiquement, mais dont les différences dans les habitudes de vie (p. ex. l'alimentation, les signaux d’accouplement) mènent à des individus génétiquement différents (spéciation sympatrique ) (chapitre 4 du volume 1 du MEA ).
Les conditions qui mènent à la spéciation font en sorte que ce processus est observable après quelques générations ou peut s’échelonner sur des périodes allant jusqu’à plusieurs millions d’années (chapitre 4 du volume 1 du MEA ![]() ; Ridley, 2009). On estime qu’il y a un peu plus de 1,6 million d’espèces recensées dans le monde (Hilton-Taylor et coll., 2009).
; Ridley, 2009). On estime qu’il y a un peu plus de 1,6 million d’espèces recensées dans le monde (Hilton-Taylor et coll., 2009).
Les analyses d’ADN ![]() et d’ARN
et d’ARN ![]() démontrent qu’il existe trois groupes d’organismes vivants : les bactéries, les archées et les eucaryotes (p. ex. les animaux, les plantes, les champignons). Un quatrième groupe, les virus, peut également être défini, bien qu’ils ne soient pas des organismes vivants tels qu’on l’entend pour les autres groupes (chapitre 4 du volume 1 du MEA
démontrent qu’il existe trois groupes d’organismes vivants : les bactéries, les archées et les eucaryotes (p. ex. les animaux, les plantes, les champignons). Un quatrième groupe, les virus, peut également être défini, bien qu’ils ne soient pas des organismes vivants tels qu’on l’entend pour les autres groupes (chapitre 4 du volume 1 du MEA ![]() ).
).
L’histoire de la Terre a été ponctuée d’épisodes importants d’extinction. Comme en fait foi la figure 5.2, jusqu’à la moitié des familles animales ont disparu au cours de ces extinctions. L’épisode le plus majeur a eu lieu il y a 250 millions d’années et il serait dû à une perturbation très importante comme de nombreuses éruptions volcaniques ou la collision d’un astéroïde avec la Terre. Le dernier épisode d’extinction a débuté il y a 30 000 à 46 000 ans selon les sources et est causé par l’action des populations humaines sur les écosystèmes.
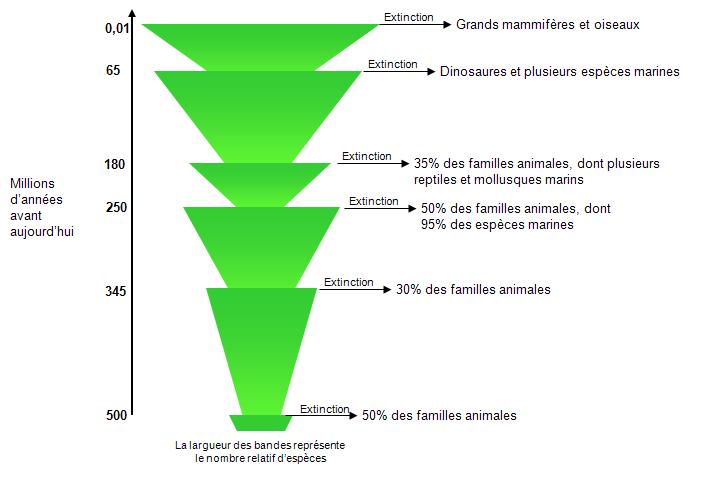
Figure 5.2 Épisodes d’extinction massive observés depuis la formation de la Terre. La largeur des bandes représente le nombre relatif d’espèces.
Source : Adapté de Primack (2008).
5.2.2 Le concept de population en écologie
Le concept de population revient constamment lorsque l’on discute de la biodiversité. Le Larousse définit ce terme comme suit :
« Ensemble des individus d'une même espèce trouvés dans une station donnée. »
Par exemple, l’aire de distribution du cerf de Virginie (Odocoileus virginianus) couvre une bonne partie de l’Amérique du Nord ainsi que l’Amérique centrale (figure 5.3). Les individus ne sont pas distribués de façon uniforme sur ce territoire, mais plutôt en groupe de plusieurs individus. Chacun de ces groupes représente une population.
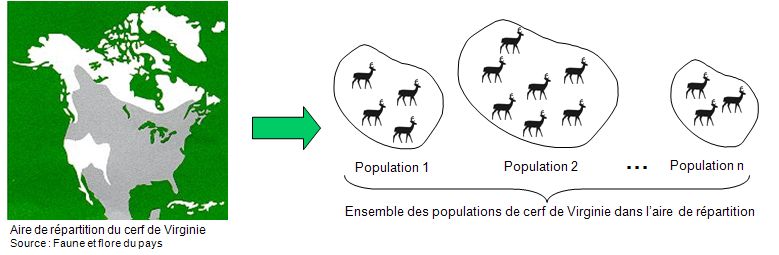
Figure 5.3 Représentation des populations de cerf de Virginie en Amérique du Nord.
Source : Adapté de Primack (2008).
5.2.3 Les fonctions dans les écosystèmes
L’évolution fait en sorte que chaque espèce joue un rôle particulier dans le fonctionnement des écosystèmes. Ces rôles peuvent être illustrés à l’aide du cycle de l’énergie (figure 5.4). Ce cycle débute par les plantes qui tirent profit d’une source d’énergie extérieure (p. ex. la lumière du soleil) ainsi que des ressources du milieu afin d’assurer leur croissance et leur reproduction. Les plantes deviennent à leur tour une source d’énergie et de nutriments pour un deuxième groupe d’êtres vivants appelé herbivore. La consommation de végétaux par ces animaux leur permet également de croître et de se reproduire. Toujours selon le cycle de l’énergie, les herbivores deviennent la proie de carnivores ou sont affectés par des parasites, permettant du même coup d’assurer le développement des individus et des populations de ces derniers organismes. Les carnivores peuvent eux-mêmes être des proies ou être parasités. Les plantes et les animaux décédés ainsi que les déchets organiques des individus vivants représentent une source d’énergie et de nutriments pour les décomposeurs. C’est grâce à ce groupe que les nutriments présents dans les individus morts peuvent retourner dans le milieu (p. ex. le sol, l'eau) pour ensuite être utilisés de nouveau par les plantes.
Ces interactions entre les espèces se déroulent dans un équilibre relatif qui est ponctué de variations attribuables entre autres aux dynamiques internes des espèces et des perturbations du milieu. Ces phénomènes ont lieu dans tous les écosystèmes connus que ceux-ci soient marins ou terrestres.
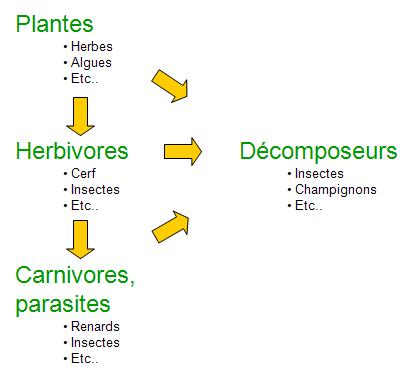
Figure 5.4 Fonctions des espèces dans les écosystèmes.
Source: Adapté de Primack (2008).
5.3 Les méthodes de mesure de la biodiversité
Étant donné la complexité des écosystèmes, la mesure de la biodiversité est un objectif difficile à atteindre. C’est pour cette raison qu’il existe de nombreux indices qui visent à mesurer l’une ou l’autre des facettes de la biodiversité. Ces indices mesurent la diversité des espèces, des gènes et des écosystèmes.
5.3.1 La diversité des espèces
À une échelle locale, la mesure la plus simple consiste à dénombrer le nombre d’espèces dans un écosystème (species richness index ![]() ). D’autres indices comme l’indice de Shannon
). D’autres indices comme l’indice de Shannon ![]() et l’indice de Simpson
et l’indice de Simpson ![]() vont plus loin en tenant également compte du nombre d’individus de chaque espèce. Il devient alors possible d’évaluer le degré d’homogénéité des espèces dans un écosystème.
vont plus loin en tenant également compte du nombre d’individus de chaque espèce. Il devient alors possible d’évaluer le degré d’homogénéité des espèces dans un écosystème.
La combinaison d’autres indices permet de connaître la diversité entre les écosystèmes. Par exemple, la diversité bêta ![]() peut être calculée en divisant le nombre d’espèces observées dans une région (diversité gamma
peut être calculée en divisant le nombre d’espèces observées dans une région (diversité gamma ![]() ) par le nombre d’espèces dans un écosystème (diversité alpha
) par le nombre d’espèces dans un écosystème (diversité alpha ![]() ) (figure 5.5) (Primack, 2004).
) (figure 5.5) (Primack, 2004).

Figure 5.5 Illustration des diversités alpha, bêta et gamma.
Source : Adapté de Primack (2008).
La diversité des espèces est également mesurable à une échelle plus large. L’indice de la Liste rouge ![]() , mis en place par l’Union internationale pour la conservation de la nature (IUCN), permet d’évaluer le degré de risque d’extinction (Hilton-Taylor et coll., 2009). Les évaluations sont habituellement réalisées à l’échelle d’une écozone, de la planète ou pour un groupe d’espèces. L’état des populations d’êtres vivants peut également être évalué grâce à l’indice Planète vivante
, mis en place par l’Union internationale pour la conservation de la nature (IUCN), permet d’évaluer le degré de risque d’extinction (Hilton-Taylor et coll., 2009). Les évaluations sont habituellement réalisées à l’échelle d’une écozone, de la planète ou pour un groupe d’espèces. L’état des populations d’êtres vivants peut également être évalué grâce à l’indice Planète vivante ![]() . Développé par le Fonds mondial pour la nature (WWF
. Développé par le Fonds mondial pour la nature (WWF ![]() ), cet indice se base sur la taille des populations, leur densité, leur abondance ou sur une estimation indirecte de leur abondance. À l’instar de l’indice de la Liste rouge, les populations sont évaluées à différentes échelles ou selon différents groupes d’espèces.
), cet indice se base sur la taille des populations, leur densité, leur abondance ou sur une estimation indirecte de leur abondance. À l’instar de l’indice de la Liste rouge, les populations sont évaluées à différentes échelles ou selon différents groupes d’espèces.
Les travaux de la Convention sur la diversité biologique ont débouché sur une approche différente de la mesure de la diversité, soit les indices sur la perte de biodiversité. Ces indices touchent l’une ou l’autre des facettes de la biodiversité et de la gestion des ressources naturelles. Les volets abordés par ces indices sont :
- Le statut des composantes de la biodiversité (p. ex. les écosystèmes, les espèces, la diversité génétique);
- L'application de la gestion durable;
- Les menaces à la biodiversité;
- L'intégrité des écosystèmes et leurs services;
- La protection des connaissances traditionnelles, des innovations et des pratiques;
- Le partage des bénéfices provenant des ressources génétiques;
- L'aide financière pour le développement.
Cette pluralité d’indicateurs permet aux décideurs gouvernementaux et non gouvernementaux d’avoir une meilleure vue d’ensemble sur la situation de la biodiversité et permet par la même occasion de faciliter la prise de décision. Vous trouverez de plus amples informations sur ces indices sur le site du Biodiversity Indicators Partnership ![]() .
.
Encadré 5.1 La modélisation des pertes et des gains en biodiversité
Les travaux de la Convention sur la diversité biologique ont également mené des groupes à modéliser les impacts des activités humaines sur la biodiversité. Dans le cas du Netherlands Environmental Assessment Agency, ces derniers utilisent des facteurs de pression environnementale comme les changements d'utilisation du territoire, les changements climatiques et les dépôts d'azote de l'atmosphère afin d'évaluer l'étendue et la biodiversité d'un écosystème. À l'instar des indicateurs présentés dans le texte, les résultats de ces modèles guident les décideurs dans leurs travaux.
5.3.2 La diversité génétique
À la base, la diversité génétique se mesure de deux façons différentes :
- Les différences observées au niveau d’un segment précis de l’ADN (locus);
- Les différences observées au niveau d’un gène.
Dans la pratique, la diversité génétique est mesurée à partir de locus marqueurs. Ces locus sont ensuite analysés afin de quantifier la variation génétique d’une population ou d’une espèce (chapitre 4 du volume 1 du MEA ![]() ).
).
La section 4.2.5 du chapitre 4 du volume 1 du MEA ![]() fournit de plus amples détails sur ce sujet. Les concepts de base de la génétique sont présentés sur le site de GENE ABC
fournit de plus amples détails sur ce sujet. Les concepts de base de la génétique sont présentés sur le site de GENE ABC ![]() du Fonds national suisse de la recherche scientifique.
du Fonds national suisse de la recherche scientifique.
5.3.3 La diversité des écosystèmes
La mesure de la diversité des écosystèmes fait l’objet d’une moins grande attention. Cette diversité est évaluée par le dénombrement d’écosystèmes identifiable sur une carte. Les cartes forestières peuvent être utilisées à cette fin, car on peut y trouver la composition, l’âge et la structure des peuplements.
- Lisez l'article de F. Le Tacon et coll. (2000) sur l'importance de la biodiversité pour le fonctionnement des écosystèmes forestiers. Accédez à cet article en cliquant sur le lien suivant :
- Lisez aussi l'article de M. A. Altieri (1999) sur l'importance de la biodiversité pour le fonctionnement des écosystèmes agricoles : Altieri, M.A. 1999. The ecological role of biodiversity in agroecosystems. Agriculture, Ecosystems and Environment 74 : 19-31. Accédez à cet article en utilisant la Réserve électronique du cours de la bibliothèque de l'Université TÉLUQ.
- De façon facultative, vous pouvez lire l'article de J.-C. Lefeuvre (1992) sur l'importance de la biodiversité pour le fonctionnement des écosystèmes agricoles. Accédez à cet article en cliquant sur le lien suivant :
5.4 Le statut de la biodiversité dans le monde
Nous avons vu précédemment que le nombre d’espèces connues sur le globe est d’un peu plus de 1,6 million d’espèces. Ces espèces ne sont pas réparties uniformément. Comme en fait foi la figure 5.6, les forêts feuillues humides tropicales et subtropicales sont de loin le biome qui abrite la plus grande diversité avec environ 20 000 espèces. La diversité des espèces dans ce biome est d’autant plus importante que près de 8000 espèces sont uniques à ce biome. Les forêts feuillues sèches tropicales et subtropicales, les prairies, les savanes et les arbustaies tropicales et subtropicales ainsi que les déserts et les arbustaies xériques représentent également une biodiversité importante avec un nombre d’espèces allant de 7 000 à 8 000 (chapitre 4 du volume 1 du MEA ![]() ).
).
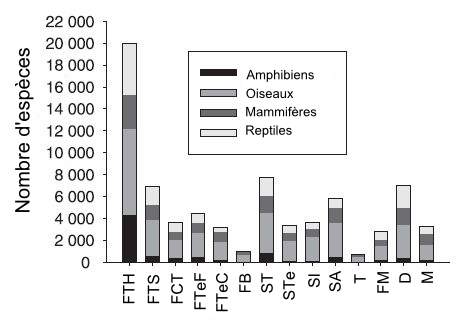
Figure 5.6 Diversité des espèces chez 14 biomes terrestres. (Les acronymes des biomes sont les suivants: FTH : forêts feuillues humides tropicales et subtropicales; FTS : forêts feuillues sèches tropicales et subtropicales; FCT : forêts conifériennes tropicales et subtropicales; FTeF : forêts tempérées feuillues et mixtes; FTeC : forêts tempérées conifériennes; FB : forêts boréales; ST : prairies, savanes et arbustaies tropicales et subtropicales; STe : prairies, savanes et arbustaies tempérées; SI : prairies et savanes innondées; SA : prairies et savanes alpines; T : toundra; FM : forêts, forêts claires et broussailles méditerranéennes; D : déserts et arbustaies xériques; M : mangrove.
Source : Adapté de Primack (2008).
Les zones de biodiversité les plus importantes et les plus en péril sont présentées dans des centres de diversité (figure 5.7). On retrouve ces centres en forêt tropicale, mais également à quelques endroits le long de la côte ouest de l’Amérique du Nord et du Sud, sur la côte est du Brésil, à la pointe sud de l’Afrique et en Océanie.
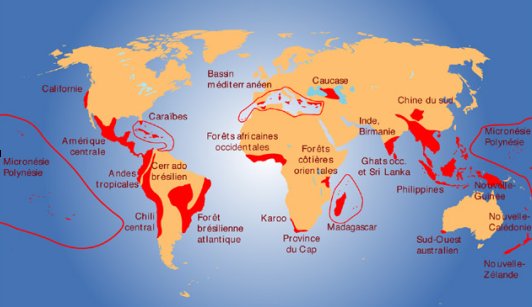
Figure 5.7 Centres de diversité dans le monde.
Source : Conservatoire botanique national de Brest. Une carte interactive, en anglais, est disponible sur le site de Conservation International ![]() .
.
Encadré 5.2 Pourquoi les forêts tropicales humides abritent-elles autant de diversité?
Trois facteurs expliquent la présence d'une telle diversité dans ces forêts :
- Les conditions climatiques : la forêt tropicale humide bénéficie de conditions climatiques qui ne limitent pas le développement des organismes. La présence continuelle d'eau et une température clémente n'obligent pas les organismes à s'ajuster à des situations qui pourraient mettre leur vie en danger (p. ex. le gel);
- La présence des arbres : la présence d'un support (p. ex. les branches) et l'ombrage créé par le feuillage favorisent des conditions de vie différentes de celles au niveau du sol. La variété des conditions de vie est favorisée par un espacement irrégulier entre les arbres et par la présence d'arbres de différentes hauteurs;
- La stabilité : les régions tropicales ont connu moins de périodes glaciaires que les autres régions. L'absence de bouleversements majeurs du milieu par la présence et la fonte de la glace a créé une stabilité qui a favorisé la spéciation.
La part la plus importante de cette diversité des espèces provient des insectes avec un peu plus de 1,2 million d’espèces, soit 75 % des espèces recensées (figure 5.8). Les plantes suivent de loin avec un peu moins de 300 000 espèces.
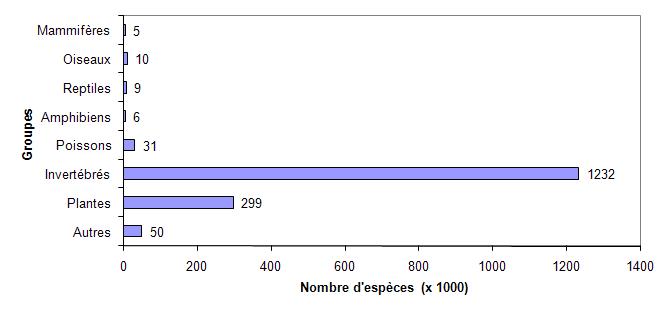
Figure 5.8 Estimation du nombre d’espèces recensées par groupe d’êtres vivants.
Source : Données tirées de Hilton-Taylor et coll. (2009).
5.5 Les impacts des activités humaines sur la diversité des espèces
Les progrès de nos civilisations ont en grande partie été possibles grâce à la conversion d’écosystèmes intacts en d’autres utilisations (p. ex. l'agriculture, l'urbanisation). Ces utilisations peuvent entraîner des transformations telles que ces écosystèmes ne sont plus en mesure de pourvoir aux besoins des organismes qui les habitaient auparavant.
5.5.1 Les causes des pertes de biodiversité
Les pertes de biodiversité résultent presque exclusivement des activités humaines. Les causes contemporaines les plus importantes sont :
- La destruction d’habitats;
- L’arrivée d’espèces envahissantes;
- Les agents pathogènes étrangers;
- La surexploitation;
- Les changements climatiques.
- Lisez la section 4.3 du chapitre 4 du volume 1 du MEA (pages 96 à 99) , laquelle décrit en détail les causes de la perte de biodiversité. Cette section fait souvent référence à l'annexe (Appendices) du volume 1 du MEA .
5.5.2 Les cas particuliers de l’agriculture et de l’exploitation forestière
Les pratiques en agriculture et en foresterie ont un impact sur la biodiversité. Ces impacts sont illustrés aux figures 5.9 et 5.10. Chacune de ces figures comprend un graphique qui présente la relation entre la biodiversité planifiée, la diversité associée et la diversité du paysage.
Dans le cas de l’agriculture, la figure 5.9 A illustre que plus le mode de culture est intensif, moins celui-ci favorise :
- La diversité planifiée. Cette diversité est celle des espèces animales et végétales exploitées ainsi que la diversité de leurs gènes. Cette plus faible diversité s’explique par la sélection effectuée depuis le début de l’agriculture, un mode de production qui favorise le plus souvent une seule espèce ainsi que par le remplacement, depuis les années 60, d’espèces adaptées localement par des variétés modernes utilisées sur de grands territoires.
- La diversité associée. On entend par diversité associée les organismes qui contribuent aux services écologiques nécessaires aux exploitations agricoles. Ces services comprennent le cycle des éléments nutritifs, le contrôle des insectes ravageurs et la pollinisation. La perte de diversité associée s’explique par le fait que l’exploitation agricole crée des conditions qui s’éloignent de celles observées dans les écosystèmes vierges. Les organismes responsables des services écologiques ne retrouvent donc pas toujours les conditions nécessaires à leur survie et au développement normal de leurs populations (p. ex. une grande gamme de périodes de floraison).
- La diversité du paysage. Cette diversité consiste en la diversité présente dans les environs des zones exploitées. La diminution de cette diversité est associée à l’homogénéisation des paysages créée par l’exploitation agricole plus intensive. Les organismes présents sur le territoire ont donc moins de possibilités d’abri et de sources de nourriture (chapitre 26 du volume 1 du MEA ).
En ce qui concerne la figure 5.9 B sur la culture de riz, il est également possible d’observer une diminution des diversités au fur et à mesure que le mode de production devient intensif. Un gain fort appréciable est cependant observable lorsque l’on abandonne l’utilisation d’herbicides et d’insecticides. Le caractère « généraliste » de ces produits diminue les populations d’insectes non ravageurs, mais également la chaîne alimentaire à laquelle ils appartiennent (chapitre 26 du volume 1 du MEA ![]() ).
).
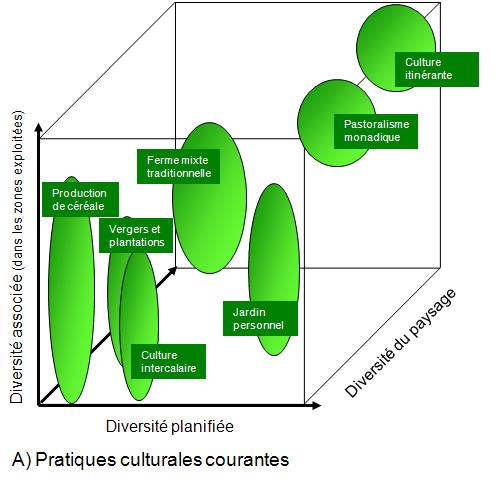
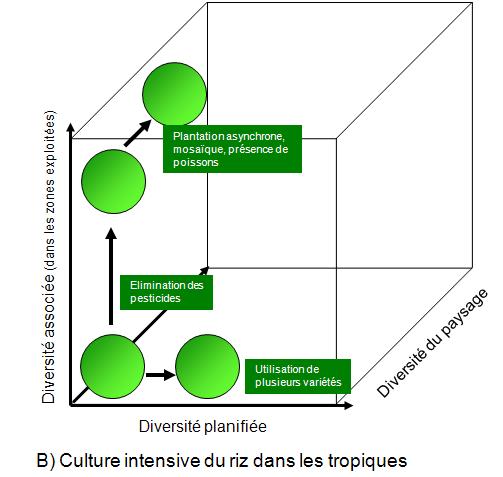
Figure 5.9 Impacts des pratiques culturales sur la biodiversité des cultures et de leur environnement immédiat. A) Pratiques culturales courantes ; B) Culture intensive du riz dans les tropiques.
Source : Adapté du chapitre 4 du volume 1 du MEA ![]() .
.
Des observations comparables sont également possibles dans le contexte forestier (figure 5.10). La culture intensive de la forêt fait en sorte que les gestionnaires forestiers sélectionnent une espèce ou un groupe restreint d’espèces afin qu’ils atteignent plus efficacement leurs objectifs de production.
L’exploitation des forêts peut également impliquer l’utilisation de pesticides et de fertilisants. Cette approche est appelée ligniculture. Certains pays comme le Canada utilisent la ligniculture sur des superficies restreintes, ce qui favorise la diversité des paysages. Ce n’est cependant pas le cas dans d’autres pays comme le Brésil où ce mode de culture est employé sur de très grands territoires.
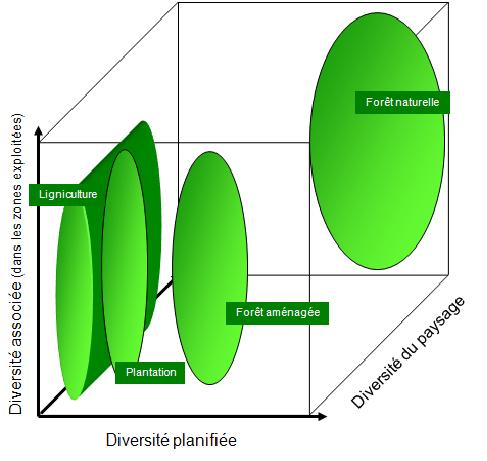
Figure 5.10 Impacts des activités forestières sur la biodiversité des forêts aménagées et de leur environnement immédiat.
5.5.3 Les pertes de biodiversité
Les activités humaines ont des impacts significatifs sur les populations animales et végétales à travers le monde. Comme en fait foi l'indice Planète vivante ![]() du Fonds mondial pour la nature, l’abondance des espèces sauvages de la planète a diminué de 30 % au cours des 35 dernières années. La figure suit l'évolution de 1 313 espèces de vertébrés, c'est-à-dire les poissons, les amphibiens, les reptiles, les oiseaux et les mammifères. On gère aussi des indices distincts pour les espèces terrestres, marines et d'eau douce.
du Fonds mondial pour la nature, l’abondance des espèces sauvages de la planète a diminué de 30 % au cours des 35 dernières années. La figure suit l'évolution de 1 313 espèces de vertébrés, c'est-à-dire les poissons, les amphibiens, les reptiles, les oiseaux et les mammifères. On gère aussi des indices distincts pour les espèces terrestres, marines et d'eau douce.
Les impacts des activités humaines diffèrent selon les régions du globe. La figure 5.11 présente, pour différentes écozones, les valeurs d’indice de la Liste rouge pour les mammifères. On remarque tout d’abord que les valeurs de l’indice diminuent au fil des années. Cette diminution signifie que le niveau de menace sur les espèces devient plus important au fil des années. On remarque également que les indices sont plus faibles dans l’écozone indomalaise. Ces faibles valeurs sont dues à l’augmentation du taux de déforestation durant les années 1990 ainsi qu’à une pression de chasse qui augmente tout particulièrement chez les espèces de taille moyenne et forte (Hilton-Taylor et coll., 2009).
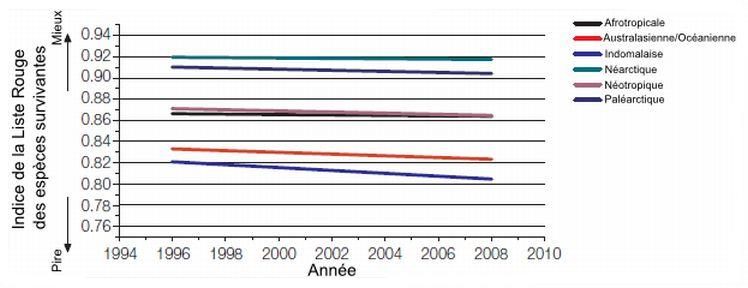
Figure 5.11 Indice de la Liste rouge des espèces de mammifères survivantes selon les écozones. (Une valeur d’indice de 1 signifie que toutes les espèces ne sont pas menacées tandis qu’une valeur de 0 montre que ces espèces seront bientôt éteintes.)
Source : Adapté de Hilton-Taylor et coll. (2009).
Lorsque l’on s’intéresse aux groupes taxonomiques les mieux connus, il est également possible d’observer une diminution des valeurs de l’indice de la Liste rouge. Les valeurs de l’indice sont plus faibles chez les amphibiens. Ces valeurs sont dues aux pertes d’habitats, la chytridiomycose (maladie fongique) et d’autres raisons qui ne sont pas encore identifiées. Le fait que ces espèces soient peu mobiles et qu’elles vivent dans des milieux limités contribuent à leur vulnérabilité. En ce qui concerne les coraux, la forte diminution observée entre les années 1995 à 2008 s’explique par les événements de blanchissement corallien observés en 1998 (Hilton-Taylor et coll., 2009).
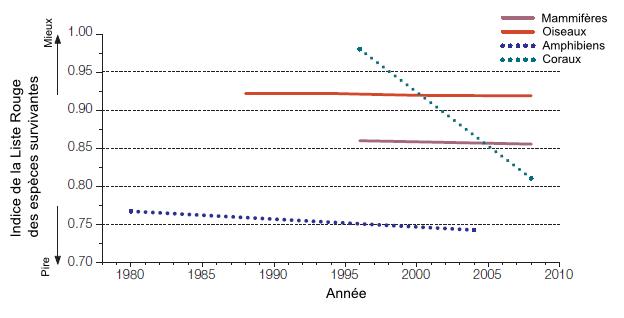
Figure 5.12 Indice de la Liste rouge pour les mammifères, les oiseaux, les amphibiens et les massifs de coraux.
Source : Adapté de Hilton-Taylor et coll. (2009).
Ces diminutions des indices se traduisent par l'extinction de plusieurs espèces alors que plusieurs d’entre elles sont en péril. Comme en fait foi le tableau 5.1, un pour cent des espèces de mammifères, soit 76 espèces, sont éteintes alors que 1219 espèces (ou un peu moins de 21 % des espèces) sont menacées de façon plus ou moins importante.
Tableau 5.1 Pourcentage1 d’espèces en péril chez les groupes taxonomiques les mieux connus.
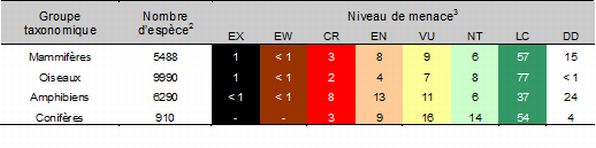
Source : Tiré de Hilton-Taylor et coll. (2009).
Légende du tableau 1 L’approche prise par l’IUCN tend à évaluer le statut des espèces en péril. Ce biais fait en sorte qu’il n’est pas possible de prendre le pourcentage d’espèces en péril (38 %) et d’affirmer que 38 % des espèces à travers le monde sont possiblement en péril (Hilton-Taylor et coll., 2009). 2 Le nombre estimé d’espèces répertoriées. 3 La classification et les codes selon la Liste rouge de l’UICN sont les suivants :
L’ensemble des activités humaines fait en sorte que le taux de perte de biodiversité atteint des niveaux jamais observés jusqu’à présent (figure 5.13). Des études des relevés fossilifères ont permis d’estimer un taux moyen d’extinction de 0,1 à 1 (une) espèce par milliers d’espèces par millénaire. Or, les extinctions observées au cours des derniers siècles mènent à des taux d’extinction de 100 à 1000 fois plus élevés que ceux observés dans les relevés fossilifères. Pour le futur, on estime que dans une centaine d’années ce taux sera 10 fois supérieur à celui que nous connaissons présentement. Une telle augmentation correspond à des taux de 1 000 à 10 000 fois plus élevés que ceux observés dans les relevés fossilifères (chapitre 4 du volume 1 du MEA ![]() ). De tels taux font en sorte que des milliers, sinon des dizaines de milliers d’espèces et des millions de populations, pourraient s’éteindre au cours des prochaines décennies (Primack, 2004).
). De tels taux font en sorte que des milliers, sinon des dizaines de milliers d’espèces et des millions de populations, pourraient s’éteindre au cours des prochaines décennies (Primack, 2004).
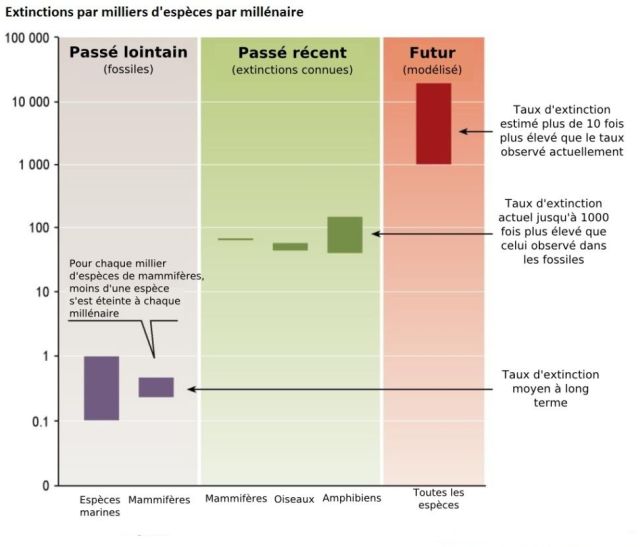
Figure 5.13 Taux d’extinction observés et estimés selon les périodes.
Source : Adapté du chapitre 4 du volume 1 du MEA ![]() .
.
Il n’est pas toujours possible de relier un événement avec la perte d’une espèce. Ceci est dû au fait que cette perte est le plus souvent observable plusieurs années après l’événement, voire une centaine d’années après celui-ci. Ce délai varie selon la durée de vie des organismes ainsi que le niveau de stress créé par l’événement.
Les délais observés avant la disparition d’espèces d’oiseaux en Amazonie varient selon la taille des fragments de forêts. Dans les fragments de moins de 100 hectares, la moitié des espèces disparut après 15 ans alors que ces délais s’échelonnent sur quelques décennies, voire peut-être sur un siècle, lorsque la taille des fragments dépasse 100 hectares (chapitre 4 du volume 1 du MEA ![]() ).
).
Les pertes de biodiversité ne sont pas l’apanage des populations contemporaines. En plus des épisodes décrits précédemment, des épisodes d’extinctions importantes ont eu lieu il y a 46 000 ans en Australie avec l’arrivée des humains sur ce continent, 12 000 ans en Amérique et il y a 2 000 ans dans le Pacifique (chapitre 4 du volume 1 du MEA ![]() ). Les causes de perte de biodiversité se concentraient principalement sur les îles océaniques et étaient liées à l’introduction d’espèces envahissantes et à la surexploitation.
). Les causes de perte de biodiversité se concentraient principalement sur les îles océaniques et étaient liées à l’introduction d’espèces envahissantes et à la surexploitation.
Les efforts de conservation portent fruit. Quarante espèces ont vu leur niveau de menace diminuer entre 2004 et 2008 (Hilton-Taylor et coll., 2009). Toutefois, ces efforts ne sont pas suffisants pour diminuer les taux de perte de biodiversité.
Les activités humaines n’ont, jusqu’en 2010 du moins, pas un effet aussi important sur la diversité génétique que celui observé sur la diversité des espèces et des biomes. Le tableau 5.2 montre que l’impact des activités humaines sur les gènes est faible (couleur jaune) par rapport aux autres échelles d’organisation écologique. En ce qui concerne la réversibilité des impacts, les niveaux sont comparables à ceux présentés dans le reste du tableau.
Tableau 5.2 Impacts des activités humaines sur la diversité génétique, des espèces et des biomes.
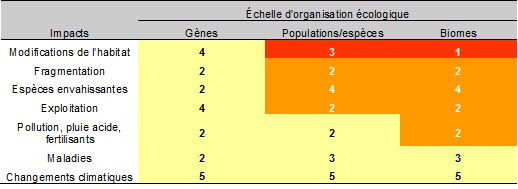
Source : Adapté du chapitre 4 du volume 1 du MEA ![]() . (Les couleurs indiquent l'importance des impacts. Rouge = maximum, Orange = moyen, Jaune = faible. Les chiffres indiquent la réversibilité des impacts. 1 facilement réversible à 5 le moins réversible.)
. (Les couleurs indiquent l'importance des impacts. Rouge = maximum, Orange = moyen, Jaune = faible. Les chiffres indiquent la réversibilité des impacts. 1 facilement réversible à 5 le moins réversible.)
L’importance des impacts des activités humaines pourrait devenir plus marquée au cours des prochaines années. La poursuite de la récolte des meilleurs individus continuera de diminuer la qualité génétique des espèces. De plus, l’augmentation de la population mondiale et du niveau de vie de plusieurs pays en développement (p. ex. la Chine, l'Inde) exerce une pression à la hausse sur les écosystèmes, ce qui entraînera la perte d’autres populations et la diminution du nombre d’individus dans les populations restantes. Ces diminutions et ces pertes prennent toute leur importance lorsque l’on réalise que la disparition d’un individu constitue une perte définitive de ses particularités génétiques et des avantages qu’elles peuvent comporter (p. ex. la capacité à s’ajuster plus facilement aux perturbations).
5.6 À partir de quel moment la perte de biodiversité devient-elle trop importante?
Il existe différentes hypothèses sur le lien qui existe entre la biodiversité et le fonctionnement des écosystèmes. Ces hypothèses sont les suivantes :
- L’hypothèse de la redondance : selon cette hypothèse, il existe une diversité minimale qui assure le fonctionnement de l’écosystème. Au-delà d’un certain seuil (à définir), les espèces qui s’ajoutent jouent un rôle similaire aux autres espèces présentes dans l’écosystème. En d’autres termes, la perte d’une espèce est compensée par les autres espèces pourvu que l’on n’ait pas atteint le seuil. Cette dépendance envers les espèces restantes augmente en contrepartie la susceptibilité de l’écosystème aux futures perturbations étant donné que chaque espèce réagit différemment aux changements de leur milieu.
- L’hypothèse du rivet : cette hypothèse associe les espèces aux rivets d’un avion. La perte d’un rivet n’entame pas la solidité de l’avion. La disparition d’autres rivets diminue graduellement la solidité de l’appareil pour mener éventuellement à son écrasement - ou à l’effondrement de l’écosystème. Cette hypothèse assume que les espèces sont relativement spécialisées et que les espèces restantes ne peuvent pas prendre le relais aussi facilement que l’hypothèse de la redondance le laisse entendre.
- L'hypothèse de la réponse idiosyncratique : cette hypothèse veut que le rôle de chaque espèce dans l’écosystème soit unique. Les impacts de la disparition d’espèces sont imprévisibles étant donné que leurs rôles sont complexes et variés;
- L'hypothèse nulle : selon cette hypothèse, la perte des espèces n’a aucun impact sur le fonctionnement normal de l’écosystème (Roselt/Oss, 2004).
La variété des écosystèmes fait en sorte qu’aucune de ces hypothèses n’explique la réalité observée sur le terrain. En pratique, les analyses révèlent que l’une ou l’autre de ces hypothèses sera dominante selon les écosystèmes et les services écologiques (Roselt/Oss 2004).
5.7 Les études de cas : comment limiter les pertes de biodiversité?
Au-delà de la protection intégrale des écosystèmes par la création d’aires protégées ou de la restauration d’écosystèmes dégradés, il est possible de limiter les pertes de biodiversité de différentes façons :
- En réalisant une agriculture ou une foresterie qui créent des conditions qui s’approchent de celles rencontrées dans les écosystèmes intacts;
- En créant des zones où l’intensité de l’exploitation est prédéfinie (p. ex. la triade au module 2 ). En créant par exemple des zones de production intensive, le gain de productivité obtenu de ces zones permet de libérer des territoires qui autrement auraient été exploités pour leurs ressources;
- En évaluant l’ensemble des impacts des mesures prises pour protéger la biodiversité. Les mesures pour favoriser la biodiversité peuvent mener à des baisses de productivité. Ces baisses peuvent mener au défrichement de territoires jusque-là riches en biodiversité (chapitre 26 du volume 1 du MEA ).
5.7.1 La gestion durable des massifs de corail
Les massifs de corail ![]() sont souvent situés dans des pays pauvres où les nombreuses ressources qu'offrent ces écosystèmes sont la plupart du temps surexploitées. Par exemple, on y exploite les poissons et les invertébrés comestibles, les poissons pour les aquariums et les coquillages pour la vente aux touristes. De plus, plusieurs groupes d'utilisateurs se servent des massifs de corail pour leurs activités de loisirs et de recherche. S'ajoutent à ces stress l'acidification des océans causée par la hausse du CO2 et le remplissage de limon par l'érosion terrestre. Maintes approches de gestion intégrée des massifs de corail ont été proposées, mais la mise en application demeure difficile compte tenu de la pauvreté des populations et de la corruption des gouvernements. Il demeure que la protection de la richesse biologique des massifs est cruciale pour plusieurs populations.
sont souvent situés dans des pays pauvres où les nombreuses ressources qu'offrent ces écosystèmes sont la plupart du temps surexploitées. Par exemple, on y exploite les poissons et les invertébrés comestibles, les poissons pour les aquariums et les coquillages pour la vente aux touristes. De plus, plusieurs groupes d'utilisateurs se servent des massifs de corail pour leurs activités de loisirs et de recherche. S'ajoutent à ces stress l'acidification des océans causée par la hausse du CO2 et le remplissage de limon par l'érosion terrestre. Maintes approches de gestion intégrée des massifs de corail ont été proposées, mais la mise en application demeure difficile compte tenu de la pauvreté des populations et de la corruption des gouvernements. Il demeure que la protection de la richesse biologique des massifs est cruciale pour plusieurs populations.
- Lisez les deux articles de T. Goreau (2009) et de Frontani et Hopkins (2008) qui analysent la complexité de la situation et les pistes de solutions pour le triangle corallien ainsi que les massifs le long des côtes du Kenya et de la Floride, respectivement. Accédez à ces articles en cliquant sur les liens suivants :
- De façon facultative, vous pouvez également visionner une vidéo sur des études du World Resource Institute portant sur la santé des massifs dans le monde. Informez-vous aussi comment leurs experts utilisent Google Earth comme outil pour recueillir des données scientifiques pour évaluer le niveau de santé des massifs en visionnant une autre vidéo .
5.7.2 La gestion de la faune aviaire en forêt par des approches sylvicoles qui imitent les perturbations naturelles
Nous venons de proposer qu'une agriculture ou une foresterie qui créent des conditions qui s’approchent de celles rencontrées dans les écosystèmes intacts puissent servir de moyen pour préserver la biodiversité. Les prochaines études en font la démonstration pour les oiseaux et les insectes en milieu forestier.
- Lisez l'article de M. J. Waterhouse et H. M. P. Armleder (2007) sur les bienfaits de la coupe partielle sur la faune aviaire de la forêt de la Colombie-Britannique, Canada. Accédez à cet article en cliquant sur le lien suivant :
- Lisez aussi l'article de P. Drapeau et coll. (2009) à propos du rôle du bois mort en forêt sur le maintien de la biodiversité aviaire au Québec : Drapeau, P. et coll. 2009. Standing deadwood for keystone bird species in the eastern boreal forest: Managing for snag dynamics. The Forestry Chronicles 85 : 227-234. Accédez à cet article en utilisant la Réserve électronique du cours de la bibliothèque de l'Université TÉLUQ.
5.7.3 La gestion de la biodiversité dans les productions agricoles
Nous avons vu au module 2 ![]() que l'agriculture s'intensifie et que cela a des conséquences sur la biodiversité des paysages agricoles. Les impacts de cette perte de biodiversité sont difficilement mesurables aujourd'hui, mais la plupart des scientifiques s'entendent pour dire que la viabilité des systèmes agricoles en sera négativement affectée à différents niveaux, selon l'environnement touché et l'utilisation du territoire. Le gouvernement anglais a été l'un des premiers à proposer des solutions pour réduire l'intensité des pratiques culturales et, de ce fait, à conserver les environnements à des niveaux se rapprochant davantage aux conditions naturelles.
que l'agriculture s'intensifie et que cela a des conséquences sur la biodiversité des paysages agricoles. Les impacts de cette perte de biodiversité sont difficilement mesurables aujourd'hui, mais la plupart des scientifiques s'entendent pour dire que la viabilité des systèmes agricoles en sera négativement affectée à différents niveaux, selon l'environnement touché et l'utilisation du territoire. Le gouvernement anglais a été l'un des premiers à proposer des solutions pour réduire l'intensité des pratiques culturales et, de ce fait, à conserver les environnements à des niveaux se rapprochant davantage aux conditions naturelles.
- Lisez l'article de G. N. Ovenden et coll. (1998) à ce sujet : Ovenden, G. N. et coll. 1998. Agri-environment scheme and their contribution to the conservation of biodiversity in England. Journal of Applied Ecology 35 : 955-960. Accédez à cet article en utilisant la Réserve électronique du cours de la bibliothèque de l'Université TÉLUQ.
Références
Hilton-Taylor, C. et coll. 2009. State of the world’s species. Dans Wildlife in a changing world – An analysis of the 2008 IUCN Red List of Threatened Species. J.-C. Vié, C. Hilton-Taylor et S. N. Stuart (éd.). IUCN, Gland, Switzerland. pages 15-41. lien ![]() .
.
Primack, R.B. 2004. A primer of conservation biology, third Edition. Sinauer Associates, Inc., Sunderland.
Primack, R.B. 2008. A primer of conservation biology, fourth Edition. Sinauer Associates, Inc., Sunderland.
Ridley, M. 2009. Modern Darwins. National Geographic 215 : 56-73.
Roselt/Oss 2004. Indicateurs écologiques - Une première approche méthodologique pour la surveillance de la biodiversité et des changements environnementaux. Réseau d'observatoires pour la surveillance écologique à long terme. Document scientifique 4. lien ![]() .
.
Spicer, J. I. 2006. Biodiversity: a beginner's guide. Oneworld Publications, Oxford.